

DIFOSFATO DE TIAMINA: atenuante del Balance Energético Negativo en Bovinos Productores de Leche.
El Balance Energético Negativo (BEN) en la vaca de alta producción lechera es una condición que se presenta invariablemente. La relación entre el ingreso-metabolismo de nutrientes y el crecimiento súbito de requerimientos es inversamente proporcional y propicia un déficit energético. El BEN se empieza a gestar desde antes del parto y se prolonga durante varias semanas posparto (7). El BEN se intensifica al inicio de la lactación por trastornos metabólicos y porque la capacidad de consumo de materia seca disminuye por causas diversas (8).
Una vaca en BEN se caracteriza por presentar las siguientes condiciones correspondientes a un Síndrome Metabólico (1) (4).

· Hipoglucemia.
· Piruvato deprimido y formación de Ácido Láctico.
· Hipoinsulinemia.
· Hormona del Crecimiento elevada.
· Gluconeogénesis.
· Cetonemia.
· Lipólisis y movilización del tejido adiposo
· Concentración elevada de Ácidos Grasos No Esterificados (AGNE).
ANÁLISIS PARTICULAR DE LOS PROCESOS METABÓLICOS, ANTES Y DURANTE EL BEN.
Hipoglucemia.
Justo antes del parto se presenta la disminución de la reserva de glucosa, glúcido que es desviado hacia el feto para terminar su desarrollo y hacia la ubre para la preparación de la leche que más tarde alimentará a la cría (7). Estas dos acciones demandan hasta el 70% de la glucosa disponible, dejando a la vaca en un estado de hipoglucemia.
La desviación de glucosa hacia feto y ubre desencadena una serie de eventos fisiológico- metabólicos que suelen ser considerados como una patología, aunque per se no lo son. Sin embargo, sus efectos si conducen a trastornos que afectan, tanto a la economía orgánica de la vaca como a su producción, reproducción y estado de salud futuros, cuando ésta se encuentre en lactación.
Los efectos de la hipoglucemia anteparto se pueden resumir básicamente en dos aspectos principales:
Cetogénesis.
Es la conversión de los aminoácidos en cuerpos cetónicos, para participar en la síntesis de moléculas de glucosa a partir de glucosa 6 fosfato.
Gluconeogénesis.
La Gluconeogénesis es inducida por la presencia de la hormona del Crecimiento (7). Es una vía emergente que se exacerba ante la depresión de la Glucólisis. La glucosa formada por esta vía, realmente no es nueva, ya que para su constitución se parte de un sustrato procedente de glucosa original formada en primera generación. Para la célula, funcionar en gluconeogénesis le implica un gasto energético alto y la mayoría de las veces no proporciona un costo beneficio grande, ya que la producción de ATP obtenida es baja (3 a 4 moléculas por revolución de Ciclo de Krebs) contra 30 obtenidos de la Glucólisis (6) (11). Además, atenta contra la síntesis de proteína y de todas las funciones emanadas de ese compuesto, como son la constitución muscular, la síntesis de hormonas y de anticuerpos. Esto cobra relevancia en los períodos de ayuno, donde los animales presentan un flujo elevado de esta vía y su capacidad de producción merma (13).
La Gluconeogénesis es una vía no deseable, sobre todo si es la única manera de que el animal esté consiguiendo glucosa.
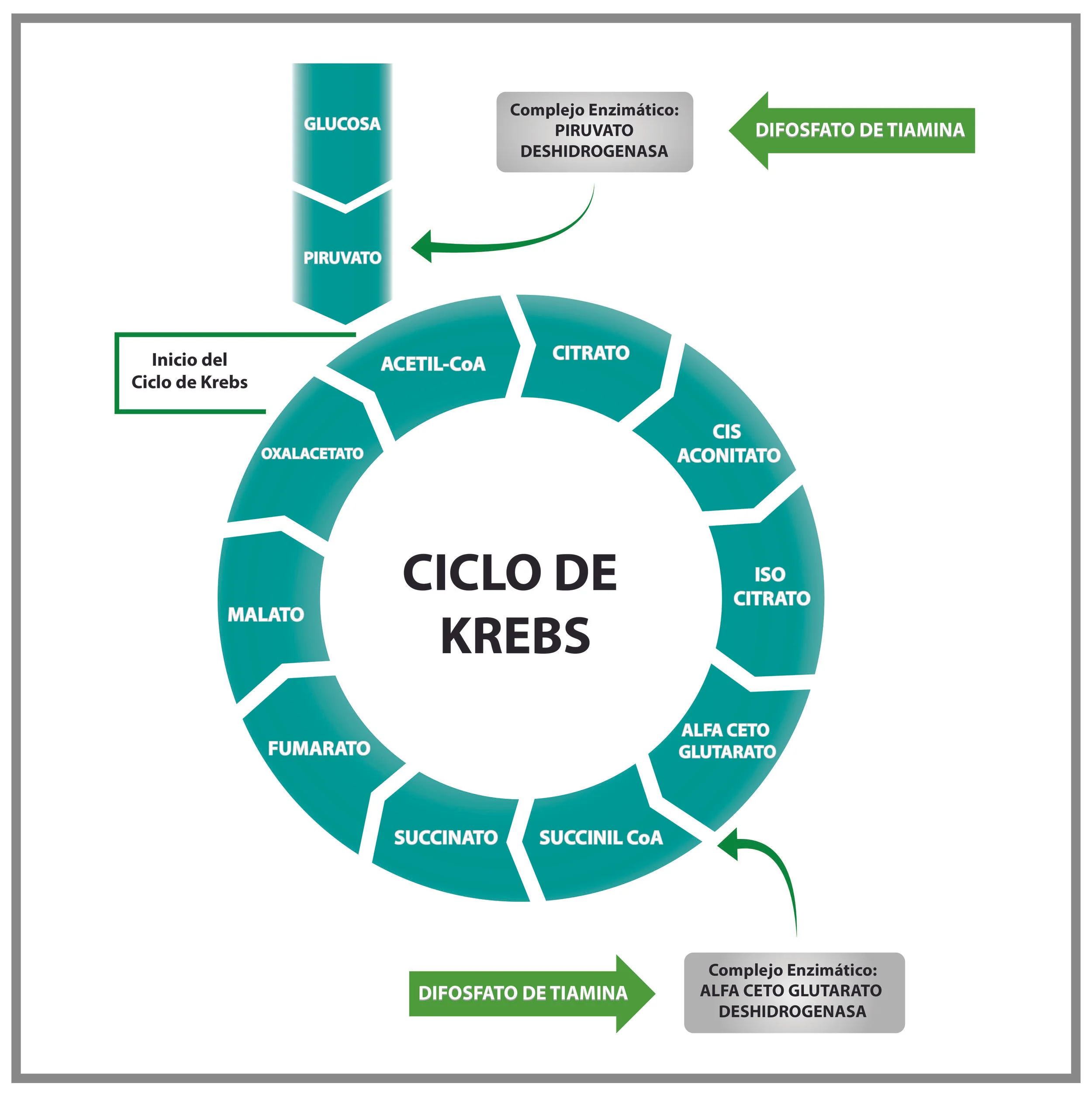
Movilización de los lípidos tisulares.
Se manifiesta de forma evidente a partir del inicio de la lactancia. Coincide con el incremento de receptores hepáticos (rHC) para la hormona del Crecimiento. La razón estriba en que siendo la grasa una reserva de energía (exclusivamente los ácidos grasos de cadena non) (13), el hígado la reconoce como una alternativa para participar en la formación de Acetil Coenzima A (producto con el que inicia su revolución el Ciclo de Krebs) (Fig.1) y representa la posibilidad de que, con su metabolismo (Beta oxidación), el ciclo productor de ATP no se detenga, aunque su costo energético sea alto se afecte al hígado (esteatosis).
Piruvato deprimido y Acidosis Láctica.
Para el inicio de cada revolución de Ciclo de Krebs es necesario que las cantidades de Acetil Coenzima A y Oxalacetato estén balanceadas, evento que no se verifica porque la producción de Piruvato (producto final de la oxidación aeróbica de la glucosa) se deprime como consecuencia de la poca cantidad de glucosa que ingresa a las células como consecuencia de la hipoglucemia e hipoinsulinemia.
REPERCUSIÓN.
El hecho de que la vaca presente hipoglucemia, cetosis, movilización de grasa y de que transite en estado gluconeogénico se traduce en los siguientes efectos negativos:
· Producción limitada de energía y no acorde a los altos requerimientos presentes en la lactancia inicial.
· Pérdida de condición corporal, que más tarde se recupera.
· Pérdida de peso vivo (proteína muscular, por su conversión a cetonas).
· Detrimento de la calidad de la leche (lactosa, grasa y proteína).
· Producción deficiente y no oportuna de las hormonas que intervienen en la reproducción, con la afectación consecuente de los parámetros reproductivos deseables.
· Producción deficiente de anticuerpos inespecíficos (inmunoglobulinas), lo que incrementa la predisposición a padecer enfermedades como mastitis e infecciones genitales.
INEXISTENTE EL ANTAGONISMO INSULINA-HORMONA DEL CRECIMIENTO.
En el argot del tema energético es común citar y aceptar como condición sine qua non el antagonismo entre las hormonas Somatotropina e Insulina presente en el BEN (10), cuando en realidad se deberían analizar por separado, ya que cada una ejerce una función específica y tiene sus propios padecimientos.
La Hipoinsulinemia no es una condición de enfermedad, ya que las células productoras de esta hormona están funcionales. Su manifestación es debida a qué las células no solicitan glucosa, porque no ven la justificación de procesar el glúcido ante la falta de requerimiento de Piruvato, por estar funcionando en Gluconeogénesis.
DIFOSFATO DE TIAMINA.
FUNCIÓN.
El Difosfato de Tiamina (Pirofosfato de Tiamina o Cocarboxilasa) es un coenzima y representa la forma proactiva de la Vitamina B1.
Sus funciones son:
· Participar en la formación de ATP dentro de los complejos I y II de la cadena respiratoria (Complejo Enzimático α-cetoglutarato deshidrogenasa y Piruvato deshidrogenasa).(5) (Fig.1)
· Participar en la descarboxilación del Piruvato y en la formación de Ácido Oxalacético, función que restablece algunos intermediarios del Ciclo de Krebs (2).
· Reducir y moderar los niveles de Gluconeogénesis.
· Propiciar una óptima utilización de glucosa de primera generación y, por ende, niveles mayores de glucógeno en hígado.
EFECTOS DE LA DEFICIENCIA DE DIFOSFATO DE TIAMINA.
Bioquímicamente, la insuficiencia de Difosfato de Tiamina se traduce en un aumento de las concentraciones de los ácidos pirúvico y láctico en la sangre y a nivel celular, del Citrato y del α-Ceto Glutarato, ya que esta coenzima interviene en las transformaciones metabólicas de dichos compuestos (3).
Además, su producción deficitaria propicia que las enzimas Piruvato Carboxilasa y AMPK se sinteticen escasamente (3).
Lo anterior es determinante para que la vaca pase de funcionar en glucogénesis atranscurrir en gluconeogénesis, ya que la Piruvato Carboxilasa es fundamental para la formación de Oxalacetato y la AMPK para que no se interrumpa la comunicación entre los tejidos y órganos que necesitan ATP y el almacén de este compuesto (3).
¿POR QUÉ ES DEFICIENTE LA PRODUCCIÓN DE DIFOSFATO DE TIAMINA EN LA VACA?

La bacteria Clostridium sporogenes es parte de la flora ruminal normal. Este microrganismo tiene la capacidad de producir una enzima llamada Tiaminasa, misma que inactiva a la Vitamina B1, escapándosele solamente pequeñas cantidades, mismas que serán suficientes para satisfacer requerimientos bajos o medios (mantenimiento de la economía orgánica y poca producción de leche) (12).
La bacteria se desarrolla mejor en condiciones de acidosis (pH <6), situación frecuente en el rumen de la vaca de alta producción y reacciona positivamente ante la presencia de Tiamina ingresada exógenamente, por lo que resulta inútil suplementar esta vitamina vía alimentación.
VENTAJAS DE ADICIONAR DIFOSFATO DE TIAMINA A LA ALIMENTACIÓN DE LA VACA ALTA PRODUCTORA.
· Estimular el Efecto Pasteur (9) al sacar rápidamente a la célula de gluconeogénesis y pasarla a glucogénesis y de esta manera restablecer la síntesis óptima de ATP y de Glucógeno, para su depósito en hígado y músculo.
· Incrementar el consumo de alimento (deseable) en beneficio al aporte de nutrientes para la producción y reproducción, al mantener constante la síntesis de glucógeno y por ende, la secreción de insulina, acción que cobra mayor relevancia durante los períodos de ayuno de la vaca (etapa postprandial o postabsortiva).
· Reducir la movilización de grasa y la pérdida de peso vivo.
· Mejorar el Pico de Lactación y obtener mejores curvas de lactación.
· Mejorar la Calidad de la Leche.
· Mejorar la Reproducción:
o Actividad ovárica precoz.
o Involuciones uterinas normales.
o Reducción de Quistes Foliculares.
o Reducción del Intervalo Parto-Primer Servicio.
o Incrementar la Fertilidad y reducir la Dosis por Concepción.
· Mantener óptimo el Sistema Inmunológico Inespecífico.
· Coadyuvar con la función energizante del hígado.
· Optimizar la utilización de los lípidos.
· Mejorar la respuesta ante los efectos del estrés.